Moser



Moser er en fellesbetegnelse som brukes om tre av rekkene i planteriket; nålkapselmoser, levermoser og bladmoser.
- Innhold
- Morfologi
- Formering
- Habitat og overlevelsesstrategier
- Biogeografiske mønster og artsdiversitet
- En verden i endring
- Les mer om rødlisting av moser i Norsk rødliste for arter 2015:
-


- Lisens
- CC BY 4.0
- Opphav
- Kristian Hassel
- Utgiver
- NTNU Vitenskapsmuseet
- Last ned
- Rediger (Krever innlogging)
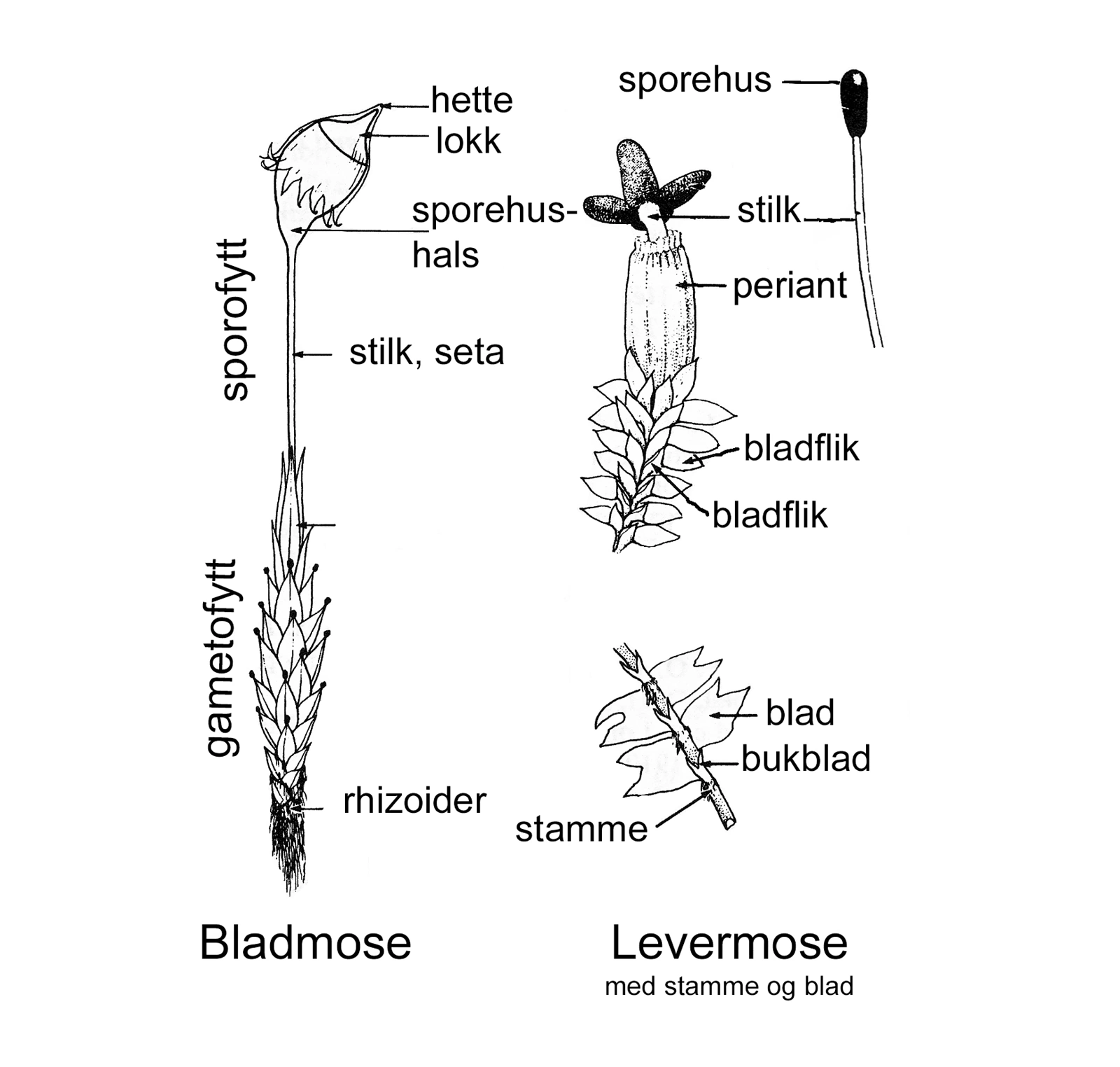
Bygning av bladmose og levermose med stengel og blad (omarbeidet etter Schofield 1988).
Moser er planter. De er ofte små, men kan i enkelte miljø stå for en vesentlig andel av primærproduksjon (fotosyntese) og artsdiversitet, slik som på myr og i fjellet. Ordet «mose» eller «mosi» kan i det norske språket bety både mosevokst og myr. Mye tyder på at det er brukt om vasstrukne mosetepper på myr. I det engelske språk brukes ordet bryophyte som stammer fra gresk og henspeiler på at planten sveller når den blir fuktig.
Morfologi
Bygningsmessig kan vi dele mosene i to hovedgrupper: de som har en enkel plantekropp med få avanserte strukturer (de thalløse) og de med stengel og blad. Alle nålkapselmoser er thalløse, blant levermosene har vi både thalløse og de med stengel og blad, mens alle bladmoser har stengel og blad. Når det gjelder antall arter, finner vi klart flest i gruppen med stengel og blad. Et typisk moseskudd består av en stengel med blader, og det kan bære en sporofytt, som er resultat av en vellykket befruktning og der sporene dannes. Moser har ikke røtter, men nesten alle har rhizoider. Dette er trådfine strukturer som vi gjerne finner på nedre del av stengelen. Noen arter har mye rhizoider og det kan da se ut som en brun filt som dekker stengelen, mens andre arter har lite eller nesten mangler rhizoider. Rhizoidene kan ha en festefunksjon og de kan gjøre vannopptak via kapillærkrefter mer effektivt. Hos noen arter kan det dannes grokorn på rhizoidene.
Vekselvåte organismer
Mosene har ingen røtter og tar hovedsakelig opp vann via overflaten. Dette kan skje indirekte ved at vann suges opp ved kapillærkrefter fra underlaget eller direkte opptak fra regnvann og luftfuktighet. Mosene må være fuktige for å drive fotosyntese, så når de tørker ut, går de inn i en hviletilstand. Mosene kan derfor sies å være vekselvåte (poikilohydriske). Et av de mest ekstreme eksemplene på dette er fjellfiltmose, som canadiske forskere fant nylig fremsmeltet fra en isbre på Ellesmere Island. Den hadde overlevd en dvaletilstand på minst 400 år.

På Ellesmere Island i Canada fant forskere fjellfiltmose Aulacomnium turgidum som våknet til live etter at den hadde vært i dvale under isen i minst 400 år.
Formering
-
- Lisens
- CC BY 4.0
- Opphav
- Kristian Hassel
- Utgiver
- NTNU Vitenskapsmuseet
- Last ned
- Rediger (Krever innlogging)
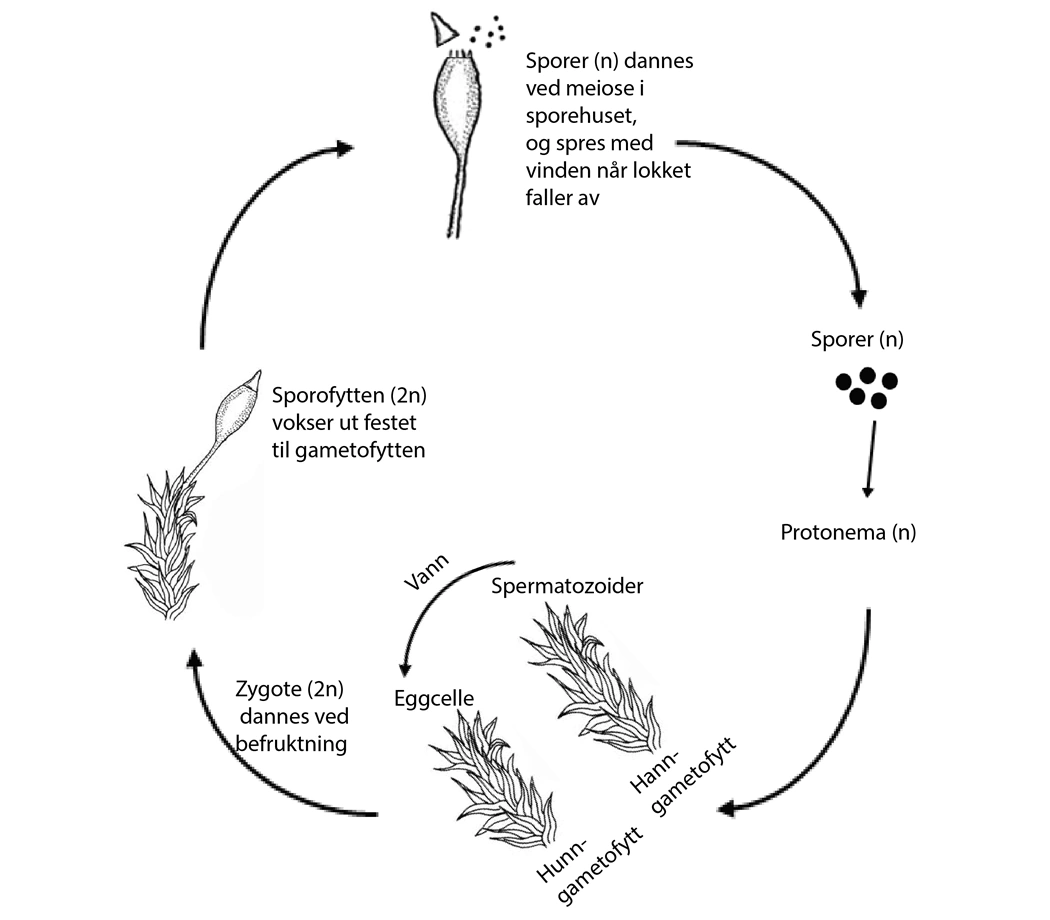
Mosenes livssyklus veksler mellom en haploid (n) gametofyttgenerasjon og en diploid (2n) sporofyttgenerasjon. Hunnlige og hannlige gameter kan produseres på det samme moseskuddet (sambu) eller de kan, som vist på figuren, sitte på ulike moseskudd (særbu). Befruktning er avhengig av at de hannlige spermatozoidene (n) overføres i vann til den hunnlige eggcellen (n), slik at en zygote (2n) dannes og sporfyttgenerasjonen kan starte sin utvikling. Sporofytten er festet på gametofytten og er ernæringmessig avhengig av denne. Sporer (n) dannes ved reduksjonsdeling (meiose) i sporofyttens sporehus. Sporene er gjerne vindspredte og en spore kan gi opphav til en ny gametofytt.
Livssyklus
Moser karakteriseres av en livssyklus med generasjonsveksling mellom en haploid og en diploid generasjon. Unikt for mosene, sammenlignet med andre landplanter, er at det er den haploide gametofyttgenerasjonen som er det dominerende livsstadiet. En annen viktig forskjell er at befruktningen er avhengig av vann.
Aseksuell formering – klonalitet og grokorn
Mange moser kan formere seg aseksuelt i tillegg til å produsere sporer. Dette er noe vi ofte ser når miljøforholdene gjør det vanskelig å gjennomføre seksuell formering, for eksempel på grunn av mangel på vann. Den enkleste formen for aseksuell formering er fragmentering. Da brekker deler av moseskuddet eller blader av og kan bli ført av sted, for eksempel med vind, vann, eller dyr. Mange moser har i tillegg mer spesialiserte strukturer for aseksuell formering som er mindre, og dermed øker sjansen for å bli spredd over større distanser. Disse strukturene kaller vi grokorn, og de kan dannes under bakken på rhizoidene, i bladhjørnene langs stengelen eller på selve bladet.
Habitat og overlevelsesstrategier
Mange moser har spesifikke krav til substratet. For eksempel foretrekker mange bustehettearter Orthotrichum spp. å vokse som epifytter på løvtrær, mens arter i gråmoseslekta Racomitrium helst vokser på sure (lav pH) bergarter. Det finnes også en del generalister, spesielt pionérarter, som stiller mindre krav til substratets egenskaper slik som vegmose Ceratodon purpureus og sølvvrangmose Bryum argenteum som vokser på bar jord.

Ospebustehette Orthotrichum gymnostomum vokser nesten bare som epifytt på osp.
Som et resultat av at ulike arter har spesifikke krav til voksestedet når det gjelder for eksempel fuktighet, pH, lys og substratets stabilitet, har det utviklet seg ulike strategier for å overleve.
Moser som lever på substrat som er kortlivede og opptrer med stor uregelmessighet, har gjerne små kortlivede gametofytter og produserer mange små sporer med god spredningsevne. Pionérarten bråtekoppmose Funaria hygrometrica er spesialist til å vokse på brannflater, et substrat som oppstår med ujevne mellomrom.
Andre moser som lever på kortlivede substrat kan ha store sporer med svært dårlig spredningsevne, og de satser da på at substratet skal oppstå igjen på omtrent samme sted. Et eksempel på en slik art er rosettgaffelmose Riccia sorocarpa.
Arter som lever i mer stabile miljøer er ofte mer langlivete og bruker det meste av sine resurser på vekst og vedlikehold, og reproduserer mer sjelden. Noen slike arter er konkurransesterke og vokser over og fortrenger andre arter. Dette gjelder for eksempel etasjemose Hylocomium splendens og storkransmose Rhytidiadelphus triquetrus som ofte kan dominere på skogbunnen.

Heigråmose Racomitrium lanuginosum tåler uttørking veldig bra, men de lyse hårspissene på bladene er en tilpasning som gjør at uttørkingen går saktere enn den ellers ville gjort.
Andre arter tåler det meste og vokser der ingen andre kan vokse. En slik stresstolerant art er heigråmose Racomitrium lanuginosum, som vokser på sure tørkeutsatte berg.
Torvmoser på myr er kanskje noen av de mest langlivede artene vi har og boreprøver viser at samme art kan ha vært tilstede på den samme myra i flere tusen år.
De mosene som har spesialiser seg mest når det gjelder å spre seg og finne veien til et nytt gunstig voksested, er artene i lemenmoseslekta Tetraplodon og møkkmoseslekta Splachnum. Som de norske navnene antyder, vokser lemenmosene gjerne på døde smågnagerkadaver og gulpeboller, mens møkkmosene vokser på ekskrementer, spesielt av hjortedyr. Dette er kortlivede substrat som kun er tilgjengelig noen få år før de forsvinner, og mosene er avhengige av effektiv og presis spredning til nye kadaver eller ekskrementer. Disse artene har nesten alltid sporofytter og produserer mange sporer, men det spesielle er at sporehusets kapselhals utskiller en lukt som tiltrekker seg tovinger som har sterk tilbøyelighet til å fly fra kadaver til kadaver eller ekskrement til ekskrement. Dermed har mosene høy sannsynlighet for å få fraktet sporene sine til et egnet voksested.

Fagerlemenmose Tetraplodon mnioides vokser vanligvis på smågnagerkadaver eller gulpeboller, men kan også, som her, vokse på reinsdyrbæsj.
Biogeografiske mønster og artsdiversitet

Trøndertorvmose Sphagnum troendelagicum er en av få endemiske moser på den Skandinaviske halvøy.
Et av de mest interessante spørsmålene relatert til mosers utbredelse i Norge er om enkelte arter kan ha overlevd på isfrie områder under siste istid eller om alle arter har innvandret etter isens tilbaketrekning. Det er sannsynligvis arter som i dag vokser i arktisk-alpine områder som kan ha overvintret siste istid her, enten på nunatakker eller isfrie områder langs kysten. Det best studerte eksemplet på dette er Andøya, hvor det er funnet flere plantefragmenter som er datert tilbake til tiden for siste istids maksimum for 24–26 000 år siden. Blant mosene i sedimentprøvene fra denne tiden er fjellfiltmose Aulacomnium turgidum og putehårstjerne Syntrichia ruralis. Disse to artene forekommer i dag sammen i kontinentale Arktiske områder, som for eksempel nordøst på Grønland. Selv om det kan finnes flere eksempler på arter som har overlevd siste istid i Norge, har de fleste innvandret etter istiden. Et argument for sterk nedising er at det finnes svært få endemiske arter i Norge og Skandinavia. De få endemiske artene vi kjenner fra disse områdene har trolig oppstått gjennom hybridisering mellom beslekta arter, slik som trøndertorvmose Sphagnum troendelagicum, som er en hybrid mellom dvergtorvmose S. tenellum og svelttorvmose S. balticum.
Innvandringshistorie, eventuell overvintring og artenes miljøkrav er viktige faktorer som bestemmer dagens utbredelse for artene. I grove trekk kan vi dele mosene inn i grupper basert på dagens utbredelse: østlige, sørlige, vestlige, arktisk-alpine og de indifferente.
-
- Lisens
- CC BY 4.0
- Opphav
- Kristian Hassel
- Utgiver
- NTNU Vitenskapsmuseet
- Last ned (486.7 KB)
- Rediger (Krever innlogging)
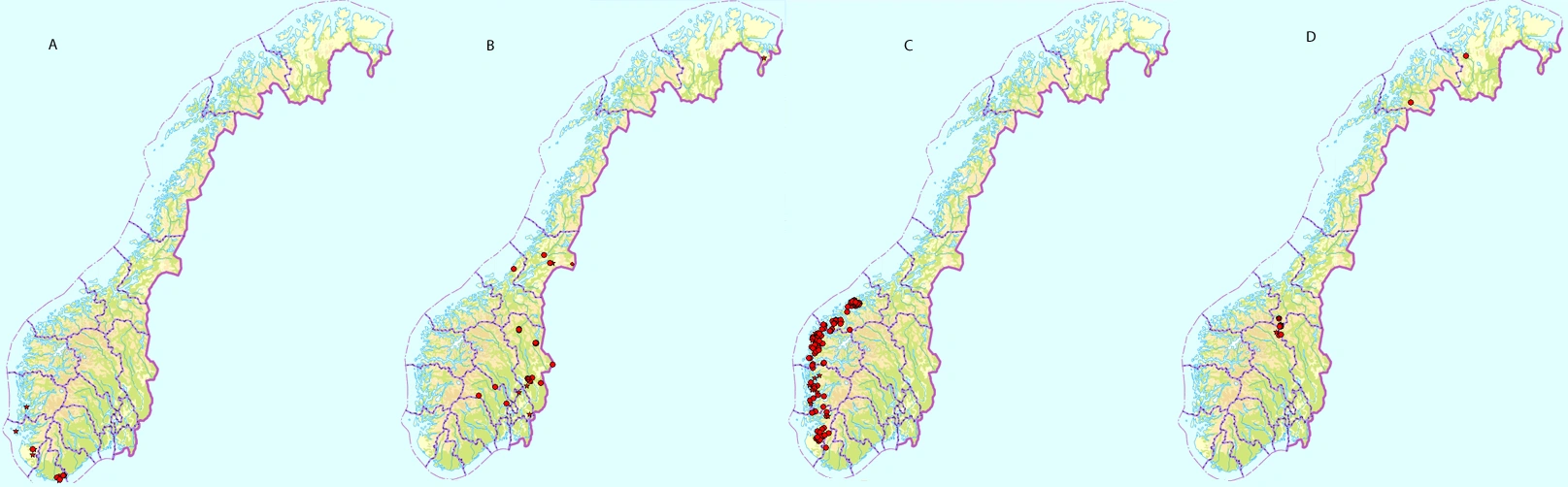
Her er eksempler på arter med de ulike utbredelsestypene. A: sørlig utbredelse hos nervesigd Dicranum fulvum, B: østlig utbredelse hos huldretorvmose Sphagnum wulfianum, C: vestlig utbredelse hos prakttvebladmose Scapania ornithopodioides og D: arktisk-alpin utbredelse hos fjellgittermose Cinclidium arcticum (kilde: http://artskart.artsdatabanken.no 2016).
En verden i endring

-
- Lisens
- CC BY 4.0
- Opphav
- Kristian Hassel
- Utgiver
- NTNU Vitenskapsmuseet
- Last ned (4.5 MB)
- Rediger (Krever innlogging)
Storbrean i Leirdalen, Jotunheimen har trukket seg tilbake flere hundre meter det siste 100-året.
Som et resultat av at tilgjengeligheten og kvaliteten på egnede leveområder stadig blir dårligere også for moser, kan det føre til en tilbakegang hos populasjonene eller at artene forsvinner fra enkelte lokaliteter. Hvert sjette år vurderes situasjonen for mosene i forbindelse med utarbeidelse av den nasjonale rødlista for arter.
Klimaendring
Effektene av temperaturøkning og økning i frekvensen av ekstremvær i form av lengre perioder med tørke er vurdert som negativ for noen av våre fjellarter. Det gjelder i første rekke arter knyttet til snøleier og andre habitater som er betinget av tilførsel av fuktighet fra snøfonner, siden størrelse og varighet av isbreer og snøfonner er i sterk tilbakegang.
Habitatødeleggelser
Fysisk ødeleggelse, kvalitetsforringelse og endringer av habitatet på grunn av menneskelige inngrep er de viktigste påvirkningsfaktorene. Det moderne bestandsskogbruket har, særlig på grunn av store flatehogster, påvirket mosefloraen negativt både direkte og indirekte. Direkte påvirkning har trolig skjedd først og fremst på grunn av endret mikroklima i områder med hogst. Drenering og grøfting av myr, sumpskog og annen våtmark til landbruks-, skogbruks-, bolig- og industriformål har hatt et betydelig omfang over lang tid. Det samme har uttak av brensel- og strøtorv. Alle disse påvirkningene, som først og fremst er konsentrert til lavlandsområdene, har negativ innvirkning på moser. Over 25 % av det opprinnelige myrarealet under skoggrensa i Norge er drenert. Vassdragsreguleringer har negativ påvirkning på moseforekomster gjennom direkte habitatødeleggelse, men påvirker også moser gjennom endrete fuktighetsforhold i overgangen mellom vann og land. Kunnskapsnivået om effekter av vassdragsregulering på moser er begrenset; særlig gjelder dette for sjeldne og truete arter. Opphør av bruk (slått og beite) og påfølgende gjengroing av semi-naturlig jordbruksmark med busker og trær påvirker også mosefloraen. En negativ utvikling er blant annet kjent for sjeldne pionérmoser med sørvestlig utbredelse knyttet til kystlynghei, som for eksempel for oddsåtemose Campylopus brevipilus og brannmose Leptodontium flexifolium.

-
- Lisens
- CC BY 4.0
- Opphav
- Kristian Hassel
- Utgiver
- NTNU Vitenskapsmuseet
- Last ned (692.2 KB)
- Rediger (Krever innlogging)
Myrområder i lavlandet har vært spesielt utsatt for ødeleggelse, som her på Heimdalsmyra i Trondheim hvor veibygging, boliger og industri i 2015 har tatt over det meste av arealet som var myr i 1957 (Kilde: Norge i bilder, https://www.norgeibilder.no/)
Referanser
Schofield WB (1985). Introduction to bryology. Macmillan Publishing Company, New York.
Schofield WB (1988). Bryophyte disjunctions in the northern hemisphere - Europe and North-America. Botanical Journal of the Linnean Society 98: 211–224.
Wickett NJ, Mirarab S, Nguyen N, Warnow T, Carpenter E, Matasci N, Ayyampalayam S, Barker MS, Burleigh JG, Gitzendanner MA, Ruhfel BR, Wafula E, Der JP, Graham SW, Mathews S, Melkonian M, Soltis DE, Soltis PS, Miles NW, Rothfels CJ, Pokorny L, Shaw AJ, DeGironimo L, Stevenson DW, Surek B, Villarreal JC, Roure B, Philippe H, dePamphilis CW, Chen T, Deyholos MK, Baucom RS, Kutchan TM, Augustin MM, Wang J, Zhang Y, Tian Z, Yan Z, Wu X, Sun X, Wong GK-S og Leebens-Mack J (2014). Phylotranscriptomic analysis of the origin and early diversification of land plants. Proceedings of the National Academy of Sciences 111: E4859–E4868. doi: 10.1073/pnas.1323926111.